一种用于提高发菜多糖产量的PGK-C-A蛋白、方法和应用
本发明属于工业微生物,尤其是一种用于提高发菜多糖产量的pgk-c-a蛋白、方法和应用。
背景技术:
1、发菜,学名发状念珠蓝细菌,属我国一级保护的陆生蓝细菌。作为一种光能自养微生物,它在生长过程中能分泌胞外多糖。微生物多糖作为天然活性大分子,不仅可以增强机体免疫力、抗肿瘤、抗炎和降血脂,还具有调节肠道菌群、提高有益菌比例以及改善花粉敏感的症状等功能,使得发菜多糖在生物医药、食品领域的潜在应用价值也日益受到人们的广泛关注。
2、微生物的胞外多糖具有可再生性、可降解性和生物安全性佳等特点。此外,蓝细菌因其自养生长、快速繁殖、强大的适应能力和可进行大规模培养等优势备受关注。目前,大多数微生物胞外多糖的商业生产依赖于异养细菌发酵,而探索新的细菌菌种则为开发新型多糖提供了一个有前景的途径。近年来,利用蓝细菌产生胞外多糖的研究引起了越来越多的关注。
3、然而,发菜多糖产量低制约着其胞外多糖大规模生产。目前,提高微生物多糖产量主要通过优化培养条件及发酵工艺、建立高通量筛选或诱变方法选择能够高效合成多糖的菌株、基因转导或基因编辑技术得到高产多糖的菌株等技术提高微生物多糖产量。例如中国发明专利cn101082057a,专利名称为:提高香菇多糖产量的高温诱导方法,此发明通过改变环境因子,对香菇液体深层发酵加以诱导,有利于香菇多糖的积累,提高香菇多糖产量,实现香菇多糖快速高效生产,相对于传统的香菇液体深层发酵,多糖产量能提高20%-30%。中国发明专利cn108456652a,专利名称:一种鞘氨醇单胞菌基因工程菌及其构建方法与应用,此发明公开了一种鞘氨醇单胞菌基因工程菌,用于发酵制备威兰胶,不外源调控ph的条件下,微生物多糖产量达到20~30g/l;用于发酵制备结冷胶时,不外源调控ph的条件下,微生物多糖产量达到15~22g/l;增强了结冷胶、威兰胶等微生物多糖的生产效率。中国发明专利cn117821346a,专利名称为:一种高产普鲁兰多糖的嗜甲烷基因工程菌及其构建方法,此发明通过在宿主菌产糖原嗜甲烷菌(alkalicoccus glycogenes)wonf2802中导入葡萄糖基转移酶ugt1和普鲁兰合酶pul1,过表达己糖激酶hk1和udpg-焦磷酸化酶udpgp1,得到高产普鲁兰多糖的嗜甲烷基因工程菌pla2。该高产普鲁兰多糖的嗜甲烷基因工程菌pla2在以甲烷或甲醇为碳源的条件下,生产得到的普鲁兰多糖产量能够达到20g/l以上。但这些技术存在成本高、效率低、难度大的缺点,且在提高发菜多糖方面不适用。
4、目前,提高发菜多糖主要采用耐氧化胁迫、优化培养方式等方法。中国发明专利cn108588143b,专利名称为:一种高效生产发菜胞外多糖的方法,此发明通过在培养过程中添加亚甲基蓝,对发菜细胞造成了氧化胁迫,而多糖更多的积累是这一胁迫的应激响应,涉及的外加亚甲基蓝的诱导方法显著提高了发菜多糖的产率,降低了成本,可用于工业规模的发酵。中国发明专利cn108531420a,专利名称为:一种同时提高发菜细胞密度和多糖产量的方法,此发明通过2,4#d诱导发菜细胞,使发菜细胞密度提高了16%同时多糖产量提高了34%,提高了发菜细胞培养的生产效率。但这些方法成本较高、对培养条件要求较高。
5、通过检索,尚未发现从pgk蛋白质的氧化翻译后修饰角度调控多糖产量的相关专利公开文献。
技术实现思路
1、本发明的目的在于克服现有技术中的不足之处,提供一种用于提高发菜多糖产量的pgk-c-a蛋白、方法和应用。
2、本发明解决其技术问题所采用的技术方案是:
3、一种通过点突变改造的用于提高发菜多糖产量的pgk-c-a突变蛋白,所述突变蛋白的氨基酸序列为seq id no.1,具体为:mskkslaslssadisgkralvrvdfnvpvddqgnitd dtriraalptiqdltqkgakvilashfgrpkgvddklrltsvakrlsellgqevvktddsigdevaakvgalqngqvlllenvrfypeeekndpefakklaanadfyvndafgtahrahastegvtkflspsvagylvekelqylqnaienpqrplaaiiggskvsskigvietllekadkliigggmiftfykarglnvgkslveedklelaksleakakergvalllptdvvladnfapdansqtvsienipdgwmgldigpdsvkffqesladtktviwngpmgvfefdkfavgteaiahtlaeigktgtttiigggdsvaavekvgladqmshistgggaslellegkvlpgiaaldda。
4、进一步地,所述pgk-c-a突变蛋白是将发菜中pgk蛋白,pgk蛋白的氨基酸序列为seq id no.2进行异源表达,pgk蛋白序列:
5、mskkslaslssadisgkralvrvdfnvpvddqgnitddtriraalptiqdltqkgakvilas
6、hfgrpkgvddklrltsvakrlsellgqevvktddsigdevaakvgalqngqvlllenvrf
7、ypeeekndpefakklaanadfyvndafgtahrahastegvtkflspsvagylvekelqyl
8、qnaienpqrplaaiiggskvsskigvietllekcdkliigggmiftfykarglnvgkslveed
9、klelaksleakakergvalllptdvvladnfapdansqtvsienipdgwmgldigpdsvkf
10、fqesladtktviwngpmgvfefdkfavgteaiahtlaeigktgtttiigggdsvaavekvgladqmshistgggaslellegkvlpgiaaldda,通过点突变将第216位半胱氨酸残基替换为丙氨酸构建版的,
11、如上所述的pgk-c-a突变蛋白的构建方法,包括如下步骤:
12、(1)发菜中pgk重组质粒的构建;
13、(2)pgk重组质粒的反向pcr得到pgk-c-a突变菌的模板质粒;
14、(3)用dpn i对模板质粒dna进行消化;
15、(4)pcr产物自身环化及转化;
16、(5)突变体pgk-c-a的确认。
17、进一步地,具体步骤如下:
18、(1)发菜基因组的提取:根据基因组提取试剂盒(vazyme,dc104-01)说明书进行提取;
19、(2)pgk基因片段的获取:以上述提取的基因组为dna模板,pgk-f、pgk-r为pgk基因的特异性引物,按下面的反应体系和程序进行pcr扩增,经琼脂糖凝胶电泳检测扩增产物后回收;
20、pgk-f的基因序列为seq id no.3,具体为:
21、aaggaaaaaagcggccgcgtgtccaaaaaaagtttagcaagtttatcttcggc;
22、pgk-r的基因序列为seq id no.4,具体为:
23、cggaattcttacgcatcatctaaagctgcaattcc;
24、pcr反应体系如下:
25、
26、pcr扩增程序如下:
27、
28、
29、(3)大肠杆菌pet-28a(+)线性化:采用notⅰ和ecorⅰ双酶切将质粒线性化,体系如下所示;通过琼脂糖凝胶电泳对酶切后的片段进行验证,将正确的片段纯化回收;随后,将纯化后的酶切片段回收产物进行测序,测序结果正确即可进行后续实验;
30、notⅰ和ecorⅰ双酶切体系:
31、
32、(4)pgk基因片段与pet-28a(+)线性化载体连接及转化
33、(a)纯化回收得到的pgk片段与载体利用t4 dna连接酶进行连接,构成重组质粒pet-28a(+)-pgk,按下面的体系混匀,于25℃金属浴2h连接,得到重组产物;
34、连接反应体系:
35、
36、(b)连接后的产物10μl于大肠杆菌dh5α感受态细胞悬液中,感受态细胞悬液中的细胞浓度为1-2×109bacteria/ml,冰浴30min,此操作于超净台中进行;
37、(c)42℃金属浴90sec,冰浴2min;
38、(d)加入1ml无抗lb液体培养基,于37℃,180rpm摇床中复苏1.5h;
39、(e)复苏后的菌液4500rpm离心2min,弃900μl上清后将下层菌体吹吸混匀,取100μl涂于在含有50μg/ml卡那霉素抗性的lb固体平板,37℃培养箱中过夜培养;
40、(5)pgk重组质粒的反向pcr得到pgk-c-a突变菌的模板质粒:
41、根据突变点设计突变引物:
42、pgk-c-a-f的基因序列为seq id no.5,具体为:gtccgccttctccagcaaagtttca;
43、pgk-c-a-r的基因序列为seq id no.6,具体为:aagctgatcatcggcggtggg;
44、按步骤(2)的反应体系和程序进行pcr扩增,反应结束后,取5μl pcr产物进行琼脂糖凝胶电泳,以确认pcr产物;
45、(6)用dpn i对模板质粒dna进行消化:pcr反应结束后,在pcr反应液中加入dpn i,每总量50μlpcr反应液中加入2μldpn i,轻轻混匀,37℃反应1小时;
46、(7)pcr产物自身环化及转化:
47、(a)从试剂盒(东洋纺,smk-101)中取出t4 polynucleotide kinase和ligationhigh,冰浴融解,融解后,将ligation high轻轻搅拌均匀;
48、(b)取一个新的pcr管,如下配制反应液:
49、pcr产物自身环化体系:
50、
51、
52、(c)轻轻搅拌均匀,16℃反应1小时;
53、(d)取部分反应液转化大肠杆菌;
54、(8)突变体pgk-c-a的确认:挑取4-8个菌落,小提质粒,采用测序进行确认。
55、如上所述的pgk-c-a突变蛋白在提高发菜多糖产量中的应用。
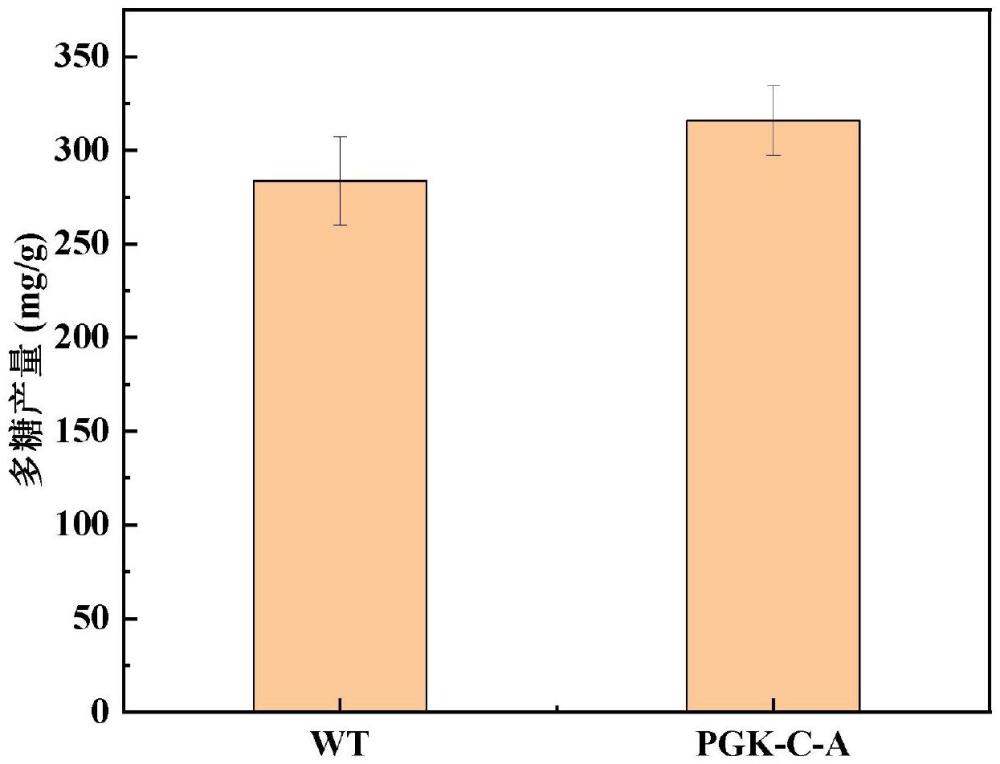
56、一种通过点突变改造pgk蛋白提高发菜多糖产量的方法,所述方法是将发菜中pgk蛋白进行异源表达,通过点突变将第216位半胱氨酸残基替换为丙氨酸构建pgk-c-a突变蛋白,pgk-c-a突变蛋白的氨基酸序列为seq id no.1,以降低pgk蛋白酶活,并采用自然转化法将其转化到发菜中,进而提高发菜多糖产量。
57、本发明取得的优点和积极效果为:
58、1、本发明通过异源表达及点突变的方法替换pgk蛋白中发生谷胱甘肽化修饰的氨基酸残基,得到一种酶活降低的pgk突变蛋白,将其命名为pgk-c-a;其酶活相对原始pgk蛋白有所降低,进而提高发菜多糖产量。
59、2、本发明方法简单可靠,操作易行,成本低廉,可用于提高发菜多糖的产量,适合大规模推广应用,对进一步构建高产多糖基因工程菌具有重要的理论和实际意义,具有广泛的应用前景。
60、3、本发明通过异源表达及点突变的方法对发菜中的pgk蛋白进行突变。pgk蛋白酶活与发菜多糖产量呈现负相关且pgk蛋白在发生谷胱甘肽化修饰后酶活升高,为降低pgk蛋白酶活,将第216位半胱氨酸残基替换为丙氨酸,从而提高发菜多糖产量。通过异源表达发菜pgk蛋白和点突变结合的方式能够降低其酶活,随后采用自然转化法将其转化到发菜菌株,进而提高发菜多糖产量,本发明方法简单,便于操作,成本低廉,可用于提高发菜多糖的产量,对进一步构建高产多糖基因工程菌具有重要的理论和实际意义,具有广泛的应用前景。
61、4、本发明从pgk蛋白质的氧化翻译后修饰角度提高发菜多糖产量,研究表明,pgk蛋白与发菜多糖产量呈现负相关且发生谷胱甘肽化修饰后pgk蛋白酶活有所增加,对发菜多糖合成起到抑制作用,因此,本发明利用点突变技术得到突变蛋白pgk-c-a,使其不发生修饰且酶活降低,以提高发菜多糖的产量,促进工业发展应用。
技术研发人员:韩培培,李梦媛,张颜,白慧芬,武佳萌,朱瑞莹
技术所有人:天津科技大学
备 注:该技术已申请专利,仅供学习研究,如用于商业用途,请联系技术所有人。
声 明 :此信息收集于网络,如果你是此专利的发明人不想本网站收录此信息请联系我们,我们会在第一时间删除